Открытие инсулина в 1921 году тесно связано с его почти немедленным клиническим применением. В 2021 году исполнилось 100 лет со дня открытия инсулина, одного из самых важных открытий в истории медицинской науки—с точки зрения его длительного воздействия на сотни миллионов людей во всем мире и развития медицинской науки.
Сто лет спустя мы узнали, что инсулин не является полностью восстановительным [6]; многие пациенты на инсулине страдают осложнениями, и необходимо строгое внимание к диетической программе.https://www.mdpi.com/1422—0067/22/3/1030/htm
Маклеод завершил свою Нобелевскую лекцию ссылкой на “запутанную проблему механизма действия инсулина в организме” [24]. Ответ был дан 50 лет спустя. Вместо того, чтобы спрашивать: “Каков механизм действия инсулина?” появление радиоиммуноанализов позволило исследователям задать вопрос: “Как клетка знает, что инсулин есть?” Была введена концепция рецепторов клеточной поверхности, обладающих способностью обнаруживать слабый сигнал на высоком фоне [25].
Рецептор инсулина представляет собой динамическую клеточную макромолекулу, которая перемещается через различные компартменты клетки на протяжении всей ее жизни. Исследования, которые проводили с использованием комбинированной молекулярной и клеточной биологии, а также биохимических методов позволили предложить следующую упорядоченную последовательность событий:
1) инсулин связывается с рецепторами, преимущественно связанными с микроворсинками на поверхности клетки; 2) инсулин запускает активацию рецепторной киназы и аутофосфорилирование, что приводит не только к инициации различных биологических сигналов, приводящих к действию инсулина, но и к перераспределению гормона-рецепторный комплекс в плоскости мембраны;
3) на не ворсинчатом домене клеточной поверхности рецепторы инсулина прикрепляются к покрытым клатрином ямкам через специфические «последовательности интернализации», присутствующие в их цитоплазматическом юкстамембранном домене;
4) комплексы рецепторов инсулина интернализуются вместе с другими рецепторами, присутствующими в тех же покрытых клатрином ямках, путем образования покрытых клатрином везикул;
5) комплексы доставляются в эндосомы, кислотный рН которых индуцирует диссоциацию молекул инсулина из рецепторы инсулина и их сортировка в разных направлениях;
6) молекулы инсулина нацелены на поздние эндосомы и лизосомы, где они деградируют;
7) рецепторы рециркулируются обратно на поверхность клетки для повторного использования.
https://pubmed.ncbi.nlm.nih.gov/2022311/
Инсулин без своего рецептора не может выполнять никаких функций [26].
Вскоре после открытия фосфорилирования тирозина [27], было продемонстрировано, что зрелый IR является тирозинкиназой [28,29,30]. Структура зрелого полноразмерного рецептора инсулина (РИ) в комплексе с инсулином была недавно решена с помощью криоэлектронной микроскопии и позволила пересмотреть модель активации РИ [32]. Примечательно, что после связывания двух молекул инсулина с перевернутыми V-образными внеклеточными альфа-субъединицами были подтвержденыдва шарнирных движения. Дальнейший интерес представляет то, что IR, по-видимому, имеет две разные конформации при низких и насыщающих концентрациях инсулина, способствуя различным активностям и предполагая, что IR может действовать как датчик циркулирующей концентрации инсулина. Различные конформационные состояния, индуцированные низкими или высокими уровнями циркулирующего инсулина, будут генерировать различные сигнальные выходы (метаболические и митогенные ответы), что означает, что уровни инсулина должны быть точно отрегулированы.
После связывания инсулина на клеточной поверхности гепатоцитов комплексы инсулина IR интернализуются и инсулин деградирует [37].
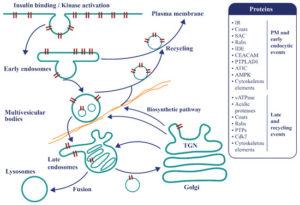
Рисунок 3. Эндосомальный аппарат представляет собой топологически универсальный тубуло-везикулярный компартмент, контролирующий сигнальные события и метаболизм. После связывания инсулина активированные тирозинкиназой комплексы инсулин-IR немедленно интернализуются в виде эндосом. В этом локусе принимается решение о рециркуляции инсулина free-IR обратно на поверхность клетки или о транспортировке активных комплексов, активируемых тирозинкиназой, к поздним компартментам для поздней сигнализации, рециркуляции или деградации. В табличке справа представлены некоторые белки, присутствующие в модуле заболевания T2D и участвующие в этих механизмах.
Недавние результаты способствуют пересмотру механизмов эндоцитоза IR и его связи с резистентностью к инсулину, а также клиренса и его связи с секрецией поджелудочной железы.
Становится понятным мнение о том, что нарушение регуляции эндоцитоза связано с резистентностью к инсулину. Интерес представляет преждевременное старение[51,52], поднимая вопрос о том, может ли IR взаимно воздействовать на передачу сигналов во время митоза.
Активность тирозинкиназы IR, по-видимому, является решающим регулятором, выбирающим лиганд-зависимые движения и передачу сигналов в эндосомах [53,54,55]. Сортировка достигается с помощью тубуловезикулярных компартментов, содержимое которых модифицируется входом и выходом везикул 70-80 нм в соответствии с моделью конверсии [56,57].
Паренхима печени является основным местом клиренса инсулина, и дефекты в этом процессе могут вызвать гиперинсулинемию и вторичную резистентность [58,59,60].
При гиперинсулинемии действие и выработка инсулина являются важными эндосомальными ответами, которые ограничивают анаболические процессы избытком питательных веществ.
Исследования дают представление о согласовании фенотипических последствий аллелей заболеваний с их генотипом и фенотипом.
В целом, результаты показывают, что причиной заболеваний, связанных с одной третью мутаций, могут быть нарушения PPINs или другие механизмы, такие как изменения локализации, а не полная потеря функции белка. ... результаты иллюстрируют силу ассоциации генотипов с фенотипами на уровне PPIN.
Модуль болезни T2D без гипотез заставляет задуматься над несколькими вопросами, касающимися неканонических инсулинзависимых механизмов, направленных на понимание первичных механизмов, с помощью которых клетки принимают решения при изменении условий окружающей среды и как изменения генотипирования отображаются на фенотипы. Важные вопросы, касающиеся модуля болезни T2D, заключаются в том, как редкие и распространенные варианты изменяют свою организацию и способствуют заболеванию и как они связаны с другими заболеваниями. Связанные вопросы — это то, как меняется архитектура во время эволюции и как эндосомы управляют консервативным гомеостатическим ответом, который может быть функциональным, избегая чрезмерного накопления питательных веществ внутри клеток в современных условиях избыточного питания.
... Cdk2 реагирует на инсулин в плазматической мембране печени и фракции Гольджи/эндосом и регулирует события слияния везикул [78]. С учетом того, что во время клеточного деления эндоцитарные события прекращаются [79] ... в стратегии “пауза для принятия решения”.
Потребность в подходах системной биологии была сформулирована клеточными биологами –“если мы не понимаем нормальную клетку, мы не можем понять болезнь”–подразумевая, что необходима помощь экспертов в различных областях наук, включая клеточную биологию, физику, математику, биомедицинские науки и клинические исследования [94].
С точки зрения вопроса Маклеода – каков механизм действия инсулина в организме? – фактический ответ обеспечивается наличием физического и функционального PPIN, где сходятся наследственные черты, также интегрирует эпигенетические предрасположенности и чувствителен к современным условиям переедания.
Актуальность темы. Механизм действия инсулина является центральной темой в биологии и медицине. В дополнение к довольно редкому состоянию дефицита инсулина, вызванному аутоиммунным разрушением β-клеток поджелудочной железы, генетические и приобретенные аномалии действия инсулина лежат в основе гораздо более распространенных состояний диабета 2 типа, ожирения и резистентности к инсулину. Последний предрасполагает к заболеваниям, начиная от гипертонии до болезни Альцгеймера и рака. Следовательно, понимание биохимических и клеточных свойств сигнализации рецептора инсулина, возможно, является приоритетом в биомедицинских исследованиях.
Передача сигналов инсулина контролирует рост клеток и метаболический гомеостаз. Инсулин оказывает комплексное воздействие на метаболизм, рост и дифференцировку клеток. Практически все клетки млекопитающих обладают IR и, следовательно, реагируют на инсулин.
Инсулин в низких дозах действует специфически через рецептор инсулина, в то время как супрафизиологические дозы инсулина действуют как через рецептор инсулина, так и через рецептор фактора роста инсулина-1. В физиологических дозах интернализованный инсулин практически не разлагается и вместо этого высвобождается неповрежденным во внеклеточную среду.https://pubmed.ncbi.nlm.nih.gov/29509435/
Рецептор инсулина (INSR) состоит как из альфа (⍺), так и из бета (β) субъединиц, где в β-субъединицах фосфорилирование тирозина более специфично для связывания с инсулином, и эта субъединица высоко экспрессируется в дифференцированной печени, мышцах и белой жировой ткани (WAT) (Wei et al., 1995). События связывания являются важным этапом для рекрутирования субстрата INSR и активации нижестоящих митогенных и метаболических сигналов (Youngren, 2007). Эти сигнальные активации зависят от концентрации инсулина. В то время как индукция метаболического ответа требует меньшего количества инсулина, митогенный ответ требует более высоких концентраций (Bedinger & Adams, 2015).
Активный IR затем интернализуется клатрин-опосредованным эндоцитозом. https://pubmed.ncbi.nlm.nih.gov/32576931/
Действие инсулина начинается со связывания с его клеточной поверхностью рецептора инсулина (IR).После связывания с инсулином активируется тирозинкиназа рецептора инсулина. Хотя активация киназы IR начинается на поверхности клетки, она поддерживается и усиливается после интернализации в эндосомальную систему (ENS) клеточной цитоплазмы.
Здравствуйте, я Раушан Турсуновна Макишева. врач-эндокринолог, кандидат мед. наук, доцент кафедры внутренних болезней Тульского государственного университета. На основе русской физиологической школы и теории функциональных систем мне удалось понять в чем причина сахарного диабета, 25 лет ушло на поиск подтверждений и практическое внедрение. На сайте публикуются статьи для пациентов, их родственников, для студентов, врачей и ученых. Это мои собственные исследования и анализ исследований, а также обзоры по наиболее актуалым и практически значимым темам.